Recherche
Des fonctions inattendues pour la dystrophine, impliquée dans la myopathie de Duchenne
Publié le 7 février 2022 – Mis à jour le 7 février 2022
Un texte de la Minute Recherche par Vincent Mirouse (iGReD, unité mixte de recherche Inserm / CNRS / Université Clermont Auvergne).
Le monde du vivant arbore une infinie diversité de formes à l’échelle des organismes entiers et cette diversité participe à notre émerveillement pour la nature. Néanmoins, l’existence de cette diversité ne repose pas sur des critères esthétiques mais bien sur des contraintes fonctionnelles. Il en va ainsi du cou de la girafe, pour atteindre les feuilles les plus hautes. La même diversité de formes, reposant sur les mêmes contraintes fonctionnelles, existe aussi à l’échelle de nos organes. Par exemple, l’arborescence de nos poumons maximise la surface d’échange entre l’air et le sang pour faciliter les échanges gazeux. Il est donc important de comprendre comment nos gènes sculptent ces organes en une forme spécifique (morphogenèse).
Les scientifiques étudient l’élongation des follicules ovariens de drosophile pour élucider les mécanismes de la morphogenèse in vivo grâce aux outils génétiques et aux possibilités d’imagerie cellulaire offerte par ce modèle animal communément utilisé dans les laboratoires. Ce follicule ovarien, précurseur du futur œuf, passe progressivement d’une forme sphérique à une forme allongée et il était connu que cette élongation dépendait des interactions entre les cellules et la matrice extracellulaire qui entoure chaque follicule. De manière inattendue les auteurs ont identifié un rôle du gène codant pour la dystrophine dans ce contexte.
Chez l’Homme, le gène de la dystrophine est muté dans les myopathies (dystrophy en anglais) de Duchenne et de Beker, maladies dégénératives à la fois fréquentes et extrêmement graves. La dystrophine appartient à un complexe transmembranaire reliant l’actine, qui constitue le cytosquelette des cellules, à la matrice extracellulaire. Il est couramment proposé que la principale fonction de la dystrophine est d’assurer une cohésion mécanique entre le cytosquelette et la matrice.
Les chercheurs ont montré chez la drosophile que la dystrophine possédait des fonctions plus complexes lors de l’élongation du follicule ovarien. D'une part, elle est nécessaire à l’organisation correcte de la matrice extracellulaire et plus particulièrement à la formation de fibrilles orientées perpendiculairement à l’axe d’élongation. D’autre part, plus tard au cours du développement du follicule, la dystrophine est aussi requise pour orienter des fibres d’actine de manière identique dans toutes les cellules. Ces fibres d’actine forment un corset moléculaire qui contraint la croissance du follicule dans un axe et provoque donc son élongation dans l’autre axe. De plus, les auteurs ont montré que les fibrilles de matrice orientées servaient de guide pour l’orientation des fibres d’actine.
Ces résultats obtenus en collaboration avec l’Université de Chicago montrent donc que la dystrophine n’agit comme une simple "colle" entre le cytosquelette d’actine et la matrice extracellulaire mais comme un organisateur de l’interface cellule/matrice. La mise en évidence de ces fonctions organisatrices dans le contexte de l’élongation du follicule ovarien pourrait permettre de mieux comprendre son rôle dans les cellules musculaires et son implication dans les myopathies.
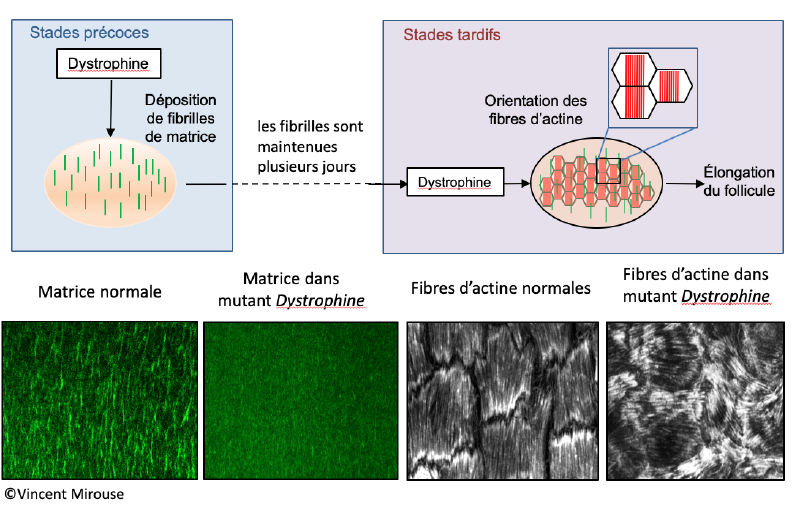
Les scientifiques étudient l’élongation des follicules ovariens de drosophile pour élucider les mécanismes de la morphogenèse in vivo grâce aux outils génétiques et aux possibilités d’imagerie cellulaire offerte par ce modèle animal communément utilisé dans les laboratoires. Ce follicule ovarien, précurseur du futur œuf, passe progressivement d’une forme sphérique à une forme allongée et il était connu que cette élongation dépendait des interactions entre les cellules et la matrice extracellulaire qui entoure chaque follicule. De manière inattendue les auteurs ont identifié un rôle du gène codant pour la dystrophine dans ce contexte.
Chez l’Homme, le gène de la dystrophine est muté dans les myopathies (dystrophy en anglais) de Duchenne et de Beker, maladies dégénératives à la fois fréquentes et extrêmement graves. La dystrophine appartient à un complexe transmembranaire reliant l’actine, qui constitue le cytosquelette des cellules, à la matrice extracellulaire. Il est couramment proposé que la principale fonction de la dystrophine est d’assurer une cohésion mécanique entre le cytosquelette et la matrice.
Les chercheurs ont montré chez la drosophile que la dystrophine possédait des fonctions plus complexes lors de l’élongation du follicule ovarien. D'une part, elle est nécessaire à l’organisation correcte de la matrice extracellulaire et plus particulièrement à la formation de fibrilles orientées perpendiculairement à l’axe d’élongation. D’autre part, plus tard au cours du développement du follicule, la dystrophine est aussi requise pour orienter des fibres d’actine de manière identique dans toutes les cellules. Ces fibres d’actine forment un corset moléculaire qui contraint la croissance du follicule dans un axe et provoque donc son élongation dans l’autre axe. De plus, les auteurs ont montré que les fibrilles de matrice orientées servaient de guide pour l’orientation des fibres d’actine.
Ces résultats obtenus en collaboration avec l’Université de Chicago montrent donc que la dystrophine n’agit comme une simple "colle" entre le cytosquelette d’actine et la matrice extracellulaire mais comme un organisateur de l’interface cellule/matrice. La mise en évidence de ces fonctions organisatrices dans le contexte de l’élongation du follicule ovarien pourrait permettre de mieux comprendre son rôle dans les cellules musculaires et son implication dans les myopathies.
Référence
Cerqueira Campos F, Dennis C, Alégot H, Fritsch C, Isabella A, Pouchin P, Bardot O, Horne-Badovinac S, Mirouse V. Oriented basement membrane fibrils provide a memory for F-actin planar polarization via the Dystrophin-Dystroglycan complex during tissue elongation. Development. 2020 Apr8;147(7):dev186957. doi: 10.1242/dev.186957. PMID: 32156755; PMCID: PMC7157587.